|
|
|
 |
|
|
The functional unit of the nervous system is the neuron, a term coined by Waldeyer in 1891. Neurons are the excitable cells of nervous tissue which conduct impulses. They arise from the neuroblasts of neural tube and neural crest origin. The complex nervous systems of vertebrates, especially humans, represent remarkably coordinated networks of these fundamental units. Thus it is not surprising to find great diversity in the form and function of individual neurons. Consequently, neurons are classified according to a number of different criteria: (1) morphology or appearance, (2) anatomical location, (3) whether they are sensory or motor, (4) conduction velocity, (5) fiber diameter, and (6) whether they are myelinated or not.
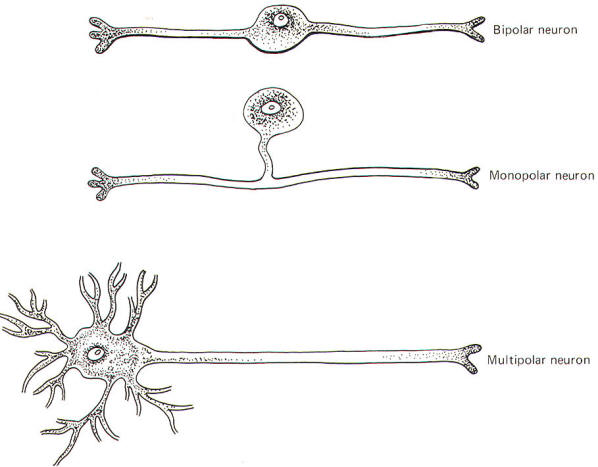
Neurons are single cells composed of a perikaryon or cell body (soma) and a variable number of neurites (processes) extending out from it. Adult neurons are classified as monopolar, bipolar, or multipolar according to whether the perikaryon has one, two, or many neurites (Fig-1).
Monopolar neurons have only one prominent neurite extending from the perikaryon, which then branches into two long processes, one central (directed toward the CNS) and one peripheral (directed away from the CNS). Most neurons of this type are sensory and are exclusively located in the peripheral nervous system. The dorsal root ganglion cells of the spinal nerves are monopolar neurons. They relay information from receptors sensitive to touch, pressure, pain, temperature, and stretch, as well as body position and movement.
Bipolar neurons have two prominent neurites extending from the perikaryon. One conducts impulses toward and one away from the soma. Bipolar neurons are found in the retina, the cochlear and vestibular ganglia, the olfactory epithelium, and in some parts of the central nervous system.
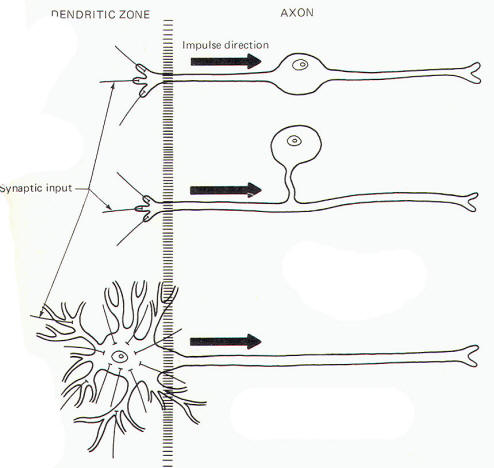
These are by far the most common type of neuron. They populate both the central and peripheral nervous systems and are characterized by several short, highly branched processes called dendrites and a single, long process extending out from the soma called an axon. A slight enlargement at the point where the axon leaves the soma, called the axon hillock, is often observed. Considerable confusion exists regarding the proper use of the terms dendrite and axon. A workable definition is that dendrites are the processes which are specialized to receive stimuli from other cells, while the axon is specialized to conduct impulses. This applies quite adequately to multipolar neurons, but the terms are arbitrary and confusing when applied to monopolar and bipolar neurons. Both the central and peripheral processes of the latter two types conduct impulses. Some neuroscientists refer to both as axons while others call the central process an axon and the peripheral process a dendrite. A system proposed by Bodian describes that portion of the neuron which is specialized to receive stimuli from other neurons or receptors as the dendritic zone. He further describes that portion which is specialized to conduct impulses (essentially the rest of the neuron) as the axon. Accordingly, the soma is included in the dendritic zone of multipolar neuron because a considerable number of synapses (contacts) from other neurons converge on it. However, no synaptic contacts are made with the somas of monopolar and bipolar neurons. In fact, only a very limited part of one of the processes of these latter two types actually receives synaptic contacts from other neurons or receptors. Thus this limited area represents the dendritic zone of monopolar and bipolar neurons. Impulses generated here are then conducted over the rest of the neuron, including the soma. In Bodian's system all of this impulse-conducting portion is the axon. The dendritic zones and axons of the three types of neurons are illustrated in Fig-2.
A neuron is afferent to a particular site if it conducts impulses toward it and efferent from that site if it conducts impulses away. For example, a neuron which conducts impulses from the thalamus to the cerebral cortex is efferent from the thalamus and afferent to the cerebral cortex. An efferent neuron which directly innervates a muscle or a gland and causes it to respond in some way is called a motor neuron. An afferent neuron which responds to changes in the external or internal environment and gives rise to conscious sensation is termed a sensory neuron. The latter is a strict definition of the term sensory. Not all afferent neurons give rise to conscious sensation, and thus not all afferent neurons are sensory. Nevertheless, the two terms (sensory and afferent) are often used interchangeably. Historically, mammalian PNS nerve fibers can be classified by group or type because of an observed correlation between conduction velocity and fiber diameter. The group system classifies afferent fibers only, while the type system classifies both. The two systems are codified in Table-1.
The long process which extends out from the soma of the nerve cell is also called a nerve fiber. These fibers are distributed throughout the peripheral nervous system in anatomically distinct structures called nerves. It is important to note that nerves exist only in the peripheral nervous system. There are no nerves within the brain or spinal cord itself. Instead, nerve fibers are distributed throughout the central nervous system in reasonably distinct anatomical groupings called nerve fiber tracts. Thus it is appropriate to speak of a spinal or cranial nerve since it is part of the peripheral nervous system but not to speak of a nerve within the brain or spinal cord. Tract is the appropriate terminology here. An example is the anterior spinothalamic tract, which is composed of a group of fibers which conduct impulses from the spinal cord to the thalamus, a route entirely within the CNS.
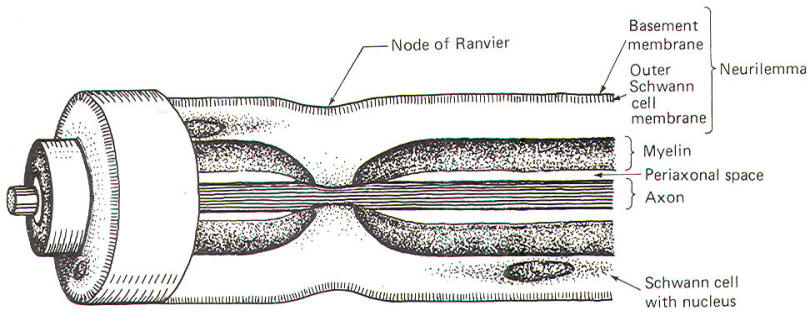
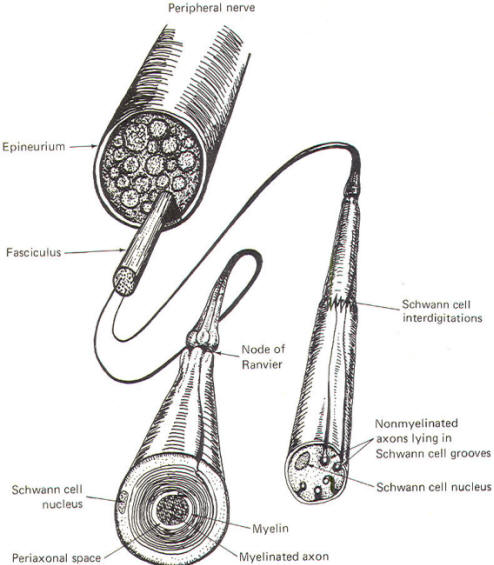
The Schwann cells are the nonexcitable cells of the peripheral nervous system. By definition they do not conduct impulses. Recall that Schwann cells are derived from Schwann cell precursors of the primitive neural crests. They develop in close association with all of the neuroblasts of the peripheral nervous system. In some cases this association is so close that the Schwann cells wrap many times around the axon of a developing neuron, laying down layer after layer of myelin and producing a myelinated neuron. In other cases, the association is not characterized by wrapping Schwann cells and the neuron remains non myelinated. Myelinated Neurons In those neurons destined to become myelinated, a Schwann cell begins to wrap around a given length of axon in a spiral fashion. In doing so, the Schwann cell extrudes its cytoplasm as its two membranes press together. In this manner it lays down layer after layer of its own membrane, forming a laminated sheath of highly lipid material called myelin. Several Schwann cells myelinate a single axon in this manner, each on a different section of its length. Because of the tight packing and lamination of the myelin, the small volume of fluid in the periaxonal space immediately surrounding the axon (Figs-3 and 4) is not readily interchangeable with the extracellular fluid of the nerve trunk. The external and internal mesaxons formed by the circling Schwann cell are not free conduits for fluid exchange. The mesaxon is the double membrane formed by the Schwann cell. Thus the axonal membrane is only in contact with a freely interchangeable fluid space at the node of Ranvier, where one Schwann cell meets another. The unique anatomical arrangement of Schwann cells around the axons of myelinated neurons endows them with a special pattern of impulse conduction called saltatory conduction. The entire Schwann cell is surrounded by a basement membrane, which together with the outer Schwann cell membrane comprises the neurilemma.
Nonmyelinated Neurons Postganglionic autonomic fibers as well as some of the very narrow diameter nerve fibers from pain and temperature receptors form a rather loose relationship with Schwann cells. These type C nerve fibers are usually found running in long, deep longitudinal depressions in Schwann cells (Fig-4). A single Schwann cell may have depressions for several narrow fibers. In this case, unlike myelinated axons, the extracellular fluid of the nerve trunk is in contact with the axonal membrane via a gap in the mesaxon which is continuous with the periaxonal space. Thus the entire non myelinated axon is in constant contact with a freely interchangeable fluid space and the pattern of impulse conduction is therefore not saltatory as observed in myelinated axons. Even though Schwann cells are in intimate contact with these axons, they are not myelinated because they have not been wrapped by the cells. The characteristics of saltatory and nonsaltatory conduction will be explained in detail elsewhere. As Fig-4 shows, Schwann cells form loose contacts with nonmyelinated axons and several of the cells interdigitate with each other enveloping the axon throughout its length.
Neuroglia ("neural glue") is a fine web of tissue which is composed of peculiar branched cells called neuroglial cells. They are located in the central nervous system only and fall into two categories: macroglia and microglia. Macroglial cells are derived from glioblasts of the neural tube and include small starshaped cells called astrocytes as well as oligodendrocytes, which are the CNS equivalent of Schwann cells. Microgliocytes are small nonneural cells, possibly of mesodermal origin. Neuroglial cells playa variety of roles in the CNS. Astrocytes appear to influence the transport of materials to the neurons of the central nervous system as well as to function to maintain an appropriate ionic environment for the neurons. Oligodendrocytes are responsible for myelinating the neurons of the central nervous system. However, unlike a single Schwann cell, which can only myelinate a single axon, each oligodendrocyte can myelinate the axons of several CNS neurons. As previously mentioned, microgliocytes are probably not of CNS origin at all. They are small cells of various forms with slender, branched processes which migrate into the CNS and act as phagocytes scavenging for waste products and breakdown components of CNS neurons.
Recall that the ependyma is a single layer of epithelial cells lining the ventricles of the brain and the central canal of the spinal cord. They arise from the fixed neuroepithelial cells lining the neural tube. Later they differentiate into the ependymal linings of the central nervous system.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
 |
||
| Fig-5 | Fig-6 |
![]() The Spinal
Cord
The Spinal
Cord
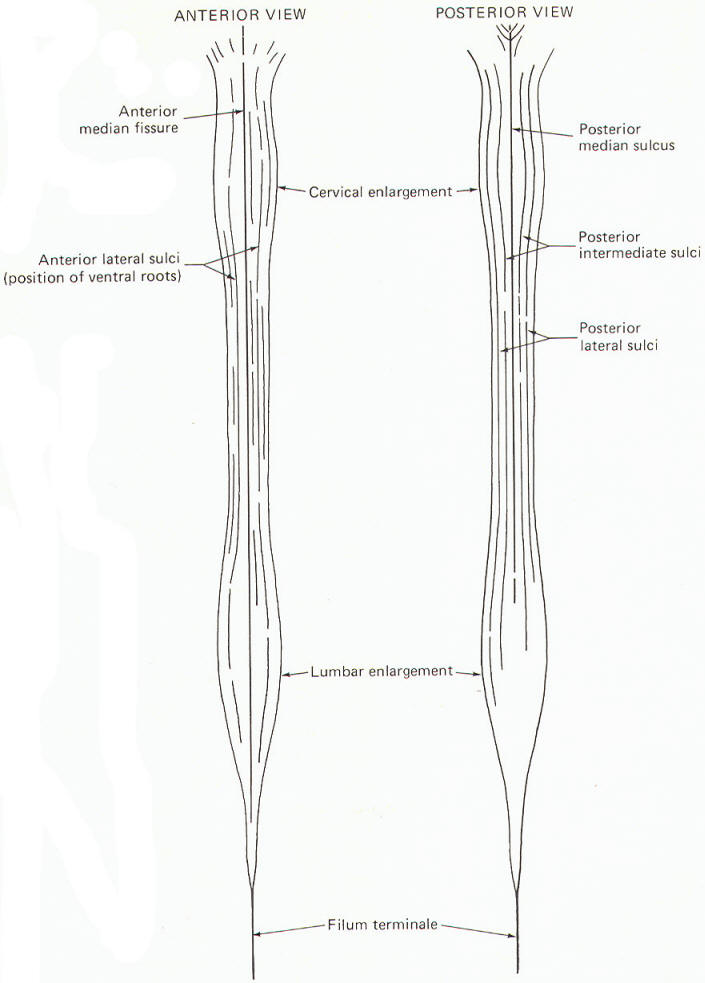
The spinal cord is the caudal extension of the brain stem into the vertebral canal. It is essentially a long, narrow structure with a cervical and lumbar enlargement. The cervical enlargement is due to the great number of afferent and efferent spinal nerve fibers from this region which innervate the arms. The lumbar enlargement represents a similar innervation of the leg musculature.
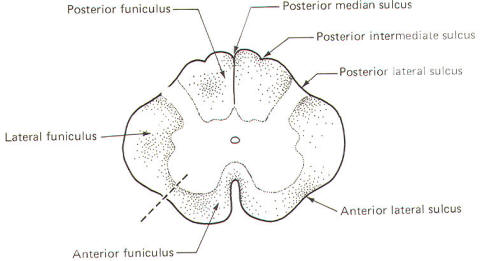
Several prominent sulci are noticeable in a posterior view of the cord (Fig-7). These include a single posterior median sulcus with posterior intermediate and posterior lateral sulci on either side of it. An anterior view shows an anterior median fissure with an anterior lateral sulcus on either side. A long, thin extension of the spinal cord, the filum terminale, extends to the coccyx at the tip of the sacrum. A cross section of the spinal cord at any level will show the characteristic butterfly-shaped pattern of gray matter surrounded by white matter. In Fig-8 notice that the relative amount of gray matter to white matter varies from one level of the cord to another.
![]() Spinal Cord
White Matter
Spinal Cord
White Matter
The spinal cord white matter is divided into three large regions called funiculi. The posterior funiculus is bounded by the posterior median and posterior lateral sulci (Fig-9). The lateral funiculus is that region of white matter between the posterior lateral and anterior lateral sulci. The anterior funiculus is bounded by the anterior lateral sulcus and the anterior median fissure. The white matter on both sides of the cord is continuous through the anterior white commissure.
 |
 |
|||
|
Fig-8 |
||||
 |
||||
|
Fig-9 |
||||
 |
||||
|
Fig-10 |
||||
 |
||||
|
Fig-7 |
Fig-11 |
![]() Ascending
and Descending Tracts in the Spinal Cord White Matter
Ascending
and Descending Tracts in the Spinal Cord White Matter
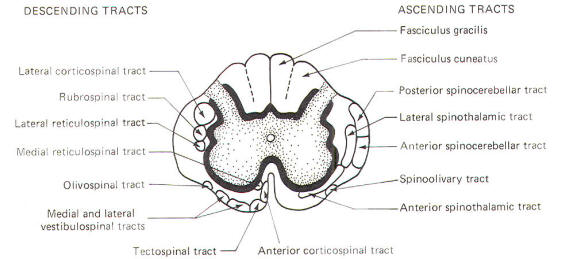
The spinal cord white matter is composed of millions of ascending and descending fibers. The ascending fibers conduct impulses up the cord while descending fibers conduct impulses downward. Most of these fibers have also been myelinated by oligodendrocytes, and it is their resulting myelin sheaths which give the white matter its characteristic color.
Most of the spinal cord fibers are grouped together in functional units called tracts. The descending tracts typically become smaller as they pass downward through the cord. This is caused by fibers continually leaving the tracts as they reach their specific destinations. Ascending tracts, with each tract associated with a functional role. The addition of a functional component as well as an examination of the clinical signs associated with selective destruction of the various tracts will make it easier to comprehend the anatomical distribution of the tracts illustrated in Fig-10.
![]() Spinal Cord
Gray Matter
Spinal Cord
Gray Matter
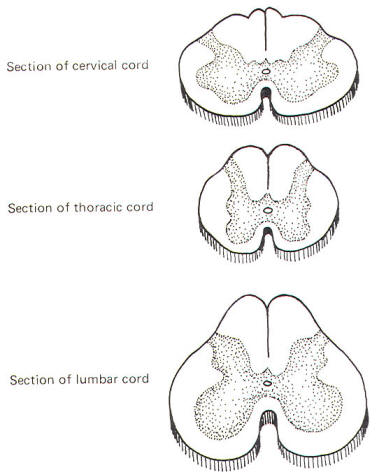
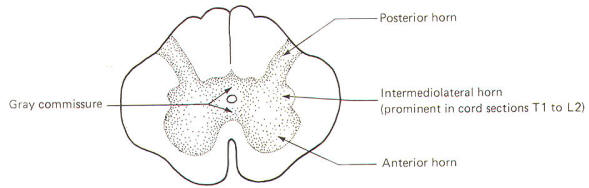
The gray matter in each half of the cord is subdivided into a posterior, intermediolateral, and anterior horn. The gray commissure connects the gray matter on each side of the cord around the central canal (Fig-11).
Recognize that Fig-11 is also a composite. Comparison with Fig-8 will help to clarify what a composite is. Because the anterior horn contains the cell bodies of motor neurons to the skeletal muscles, it is considerably larger in the cervical and lumbar enlargements where the cord gives rise to the spinal nerves innervating the arms and legs. Also, because most of the sensory fibers of spinal nerves terminate in the posterior horn, it is not surprising to find a larger horn in the cervical and lumbar regions than in the thoracic cord.
An intermediolateral horn, which gives rise to preganglionic sympathetic neurons, is found only in cross sections of the cord between T1 and L2. A similar region giving rise to preganglionic parasympathetic neurons is located in the intermediate gray matter of sacral cord segments 2 to 4. However, unlike segments T1 through L2, it does not extend as a noticeable lateral "horn."
![]() Laminar
Architecture of the Gray Matter
Laminar
Architecture of the Gray Matter
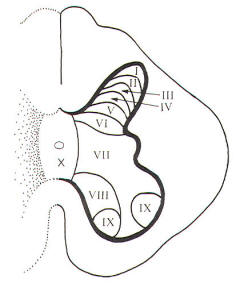
A convenient way to subdivide the gray matter of the spinal cord is according to the general cytoarchitecture found in its various regions. These cell regions, or laminae, are illustrated in Fig-12. It should be noted that this scheme is based on the spinal cord of the cat. Nevertheless, the system is being applied with due caution to humans.
 |
| Fig-12 |
Laminae I, II, III, and IV are thought to be the principal sensory receiving areas for afferent input to the cord. Laminae V and VI deal with proprioceptive input (dealing with body position and movement) as well as input from the cerebral cortex and other higher centers. Lamina VII has connections with many higher centers. Lamina VIII receives input from the opposite side of the cord as well as having numerous connections with higher brain centers. Lamina IX is the region of alpha and gamma motor neurons to skeletal muscles. Lamina X is probably a commissural area. It cannot be overstressed that the suspected roles assigned to the laminae above represent a considerable oversimplification. Nevertheless, it gives a basis for understanding synaptic relays as they relate to ascending and descending tracts in the cord and afferent input and efferent output with spinal nerves.
![]() PERIPHERAL
NERVOUS SYSTEM
PERIPHERAL
NERVOUS SYSTEM
The peripheral nervous system is composed of 12 pairs of cranial nerves and 31 pairs of spinal nerves. It represents an extension of the central nervous system into the far reaches of the body. Each spinal nerve contains both afferent and efferent fibers while cranial nerves, on the other hand, are more diverse. Some are afferent only, some are efferent only, and some are mixed (both afferent and efferent).
![]() Spinal
Nerves
Spinal
Nerves
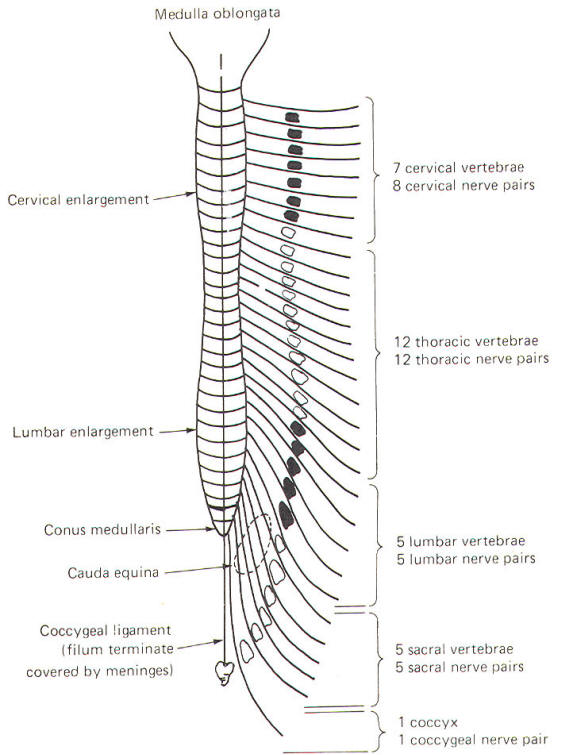
Each pair of spinal nerves extends laterally from the cord at regular intervals from cervical to coccygeal regions. The spinal nerves leave the vertebral canal via regular openings in the vertebral column called intervertebral foramina (Fig-14). Each spinal nerve is named according to the intervertebral foramina through which it exits. Notice in Fig-13 that there is one spinal nerve pair for each vertebra with the exception of the cervical region, where there are seven cervical vertebrae but eight cervical nerve pairs.
 |
 |
| Fig-13 | Fig-14 |
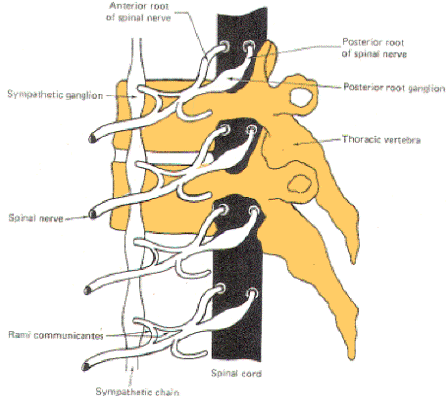
Each spinal nerve communicates with the spinal cord via two short roots, one posterior (dorsal) and one anterior (ventral). The anterior and posterior roots of the spinal nerves lie entirely within the vertebral canal. They join and form the spinal nerve just before it exits the vertebral canal through the intervertebral foramen.
At the point where spinal nerves T1 through L2 leave their respective foramina, two short arms (rami communicantes) connect the spinal nerve with a sympathetic ganglion lying adjacent to the body of the vertebra (Fig-14). A ganglion is a group of cell bodies located outside of the CNS. These 14 ganglia are connected to each other to form a vertical sympathetic chain. Three additional ganglia, the superior, middle, and inferior cervical, join the chain superiorly while another three to five ganglia in the lumbar region join it inferiorly. Thus 20 to 22 ganglia make up the sympathetic chain on each side of the cord. The three cervical ganglia and the last three to five ganglia in the chain communicate with spinal nerves via only one ramus while the ganglia associated with spinal nerves T1 through L2 communicate through two.
![]() Cranial
Nerves
Cranial
Nerves
There are 12 pairs of cranial nerves communicating directly with the brain. They are analogous with the 3 1 pairs of spinal nerves which communicate with the spinal cord. They include the following:
|
I |
Olfactory |
VII |
Facial |
|
II |
Optic |
VIII |
Vestibulocochlear |
|
III |
Oculomotor |
IX |
Glossopharyngeal |
|
IV |
Trochlear |
X |
Vagus |
|
V |
Trigeminal |
XI |
Accessory |
|
VI |
AbducCNS |
XII |
Hypoglossal |
![]() Cranial
Nerve Emergence and the Brainstem
Cranial
Nerve Emergence and the Brainstem
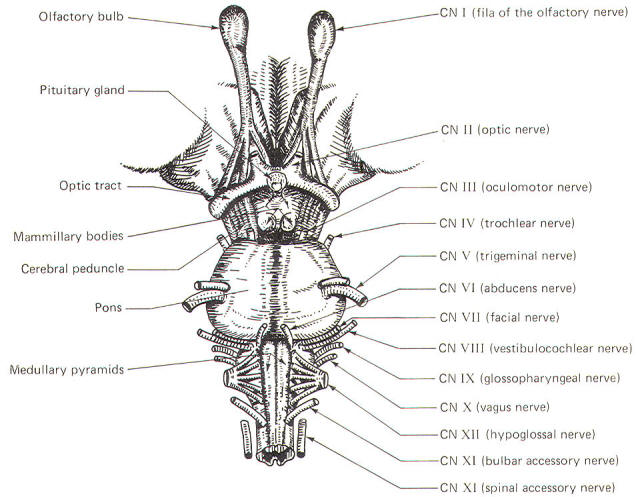
The cranial nerves and their relationship to the brain stem are shown in Fig-15. The olfactory nerves (I) enter through the cribiform plate of the ethmoid bone as 20 to 30 fila (threadlike structures) to contact the olfactory bulb. The optic nerves (II) cross anterior to the pituitary gland in the optic chiasm and continue around the cerebral peduncles of the midbrain as the optic tracts. The oculomotor nerves (III) originate in the midbrain and emerge anteriorly close together at the superior border of the pons. The trochlear nerves (IV) emerge from the posterior surface of the midbrain just below the inferior colliculi. From here they wrap around the cerebral peduncles to appear anteriorly at the superior border of the pons. The trigeminal nerves (V) arise from the anterolateral surface of the pons. The abducens nerves (VI) originate in the pons and emerge close together at the anterior inferior border of the pons. The facial nerves (VII). originating in the pons. and the vestibulocochlear nerves (VIII), which originate in the upper medulla. emerge laterally at the pontomedullary border.
 |
|
||
| Fig-15 |
Emerging from the lateral medulla posterior to the olive (a rounded elevation lateral to the pyramids in the medulla oblongata) in order from superior to inferior are the glossopharyngeal nerves (IX), vagus nerves (X), and bulbar accessory nerves (XI). Appearing with them but originating in the spinal accessory nuclei of the upper cervical cord are the spinal portions of the accessory nerves. the spinal accessory nerves (XI). Finally, arising from the lateral medulla anterior to the olive are the hypoglossal nerves (XII).
![]() CLASSIFICATION OF SPINAL AND CRANIAL NERVE FIBERS
CLASSIFICATION OF SPINAL AND CRANIAL NERVE FIBERS
Spinal nerve fibers are either afferent or efferent with respect to the spinal cord (Table-2). The spinal efferent fibers are either somatic (innervating skeletal muscles derived from mesodermal somites) or visceral (innervating cardiac muscle, smooth muscle, or glands). Similarly, spinal afferent fibers are also classified as either somatic or visceral. All are called general fibers.
Cranial nerve fibers are classified in the same manner as spinal nerve fibers (Table-2). However, there is also a special classification. Nerve fibers classified as special innervate the special sense organs involved in hearing, seeing, smelling, and tasting. In addition, special fibers innervate the vestibular system and those skeletal muscles derived from the mesoderm of the branchial arches (embryonic segments which give rise to structures in the ear and neck).
| Table-2 Classification of Spinal and Cranial Nerve Fibers | ||||
| I | Spinal nerve fiber classification | |||
| A | General afferent fibers. | The afferent unipolar neurons of the posterior root of the spinal cord with cell bodies in the posterior root ganglia | ||
| 1 | General somatic afferent (GSA) | From exteroceptors responding to touch, pressure, pain, and temperature as well as from the proprioceptors of muscles, tendons, and joints | ||
| 2 | General visceral afferent (GVA) | From interoceptors of the viscera | ||
| B |
General efferent fibers. |
The efferent multipolar neurons with cell bodies in the anterior and intermediolateral horns of the spinal cord gray matter | ||
| 1 | General somatic efferent (GSE) | The alpha and gamma motor neurons to somatic skeletal muscle and muscle spindles with cell bodies in the anterior horn of the spinal cord gray matter | ||
| 2 | General visceral efferent (GVE) | The autonomic fibers to cardiac muscle, smooth muscle, and glands | ||
| II | Cranial nerve fiber classification | |||
| A | General afferent fibers. | The afferent unipolar neurons with cell bodies in the craniospinal ganglia | ||
| 1 | General somatic afferent (GSA). | From exteroceptors responding to touch, pressure, pain, and temperature as well as from the proprioceptors of muscles, tendons, and joints | ||
| 2 | General visceral afferent (GVA). | From interoceptors of the viscera | ||
| B | Special afferent fibers. | The afferent neurons from the special sense organs (eye, ear, nose, and tongue) and the vestibular system | ||
| 1 | Special somatic afferent (SSA). | Exteroceptors from the eye and ear as well as proprioceptors from the vestibular system | ||
| 2 | Special visceral afferent (SVA). | Exteroceptors from the olfactory epithelium and the taste buds | ||
| C | General efferent fibers. | The efferent neurons originating in brainstem nuclei innervating somatic skeletal muscle as well as those innervating cardiac muscle, smooth muscle, and glands | ||
| 1 | General somatic efferent (GSE). | To somatic skeletal muscles | ||
| 2 | General visceral efferent (GVE). | The autonomic fibers to cardiac muscle, smooth muscle, and glands | ||
| D | Special efferent fibers. | The efferent neurons originating in brainstem nuclei innervating branchiomeric skeletal muscle | ||
| 1 | Special visceral efferent (SVE). | To branchiomeric skeletal muscles | ||
|
|
||
|

Copyright [2007] [CNS Clinic-Jordan]. All rights reserved